CRES-T法とは?
植物には機能が重複した転写因子が数多く存在し、一つの遺伝子の欠損では同等の機能を持つ別の転写因子が補償的に働き、表現型に影響が表れないことがしばしばある(図1)。
転写因子にリプレッションドメインを付加したキメラリプレッサーを植物に導入すると、内在性の転写因子だけでなく機能重複する転写因子の転写活性能に優先して標的遺伝子の発現を抑制することから、その転写因子の欠損株と同様な表現型が表れる(図2)。このようなキメラリプレッサーによる遺伝子機能サイレンシング技法を
C
himeric
re
pressor
s
ilencing
t
echnology (CRES-T法)と呼び、植物転写因子の機能を解明する新しい手法として注目されている。
図1
図2
例えば、シロイヌナズナのAP3、AGは花器官形成に働く転写因子であり、
ap3
変異体では花弁とおしべが欠失、
ag
変異体では八重咲きの花となる。AP3、AGにリプレッションドメインを付加したキメラリプレッサーを植物体内で発現させると、それぞれの変異体と同様の表現型が得られた(図3、4)。また、茎頂分裂組織で働く
CUC1
、
CUC2
は、それぞれ単独の変異体では表現型が表れないが、
cuc1 cuc2
二重変異体にすると子葉が融合しカップ状になる。CUC1にCRES-T法を適用した形質転換体では、カップ状の子葉形成が認められ、CUC2の場合でも同様の結果であった(図5)。このことはCRES-T法が遺伝子重複をも克服してドミナントに作用することを示しており、CRES-T法の優れた点の1つとなっている。
図3
左:野生型
右:AP3キメラリプレッサー発現体
図4
左:野生型
右:AGキメラリプレッサー発現体
図5
左:野生型
右:CUC2キメラリプレッサー発現体
参考文献
Hiratsu K, Matsui K, Koyama T, Ohme-Takagi M. (2003)
Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in
Arabidopsis
.
Plant J. 34: 733-9. (
PubMed
)
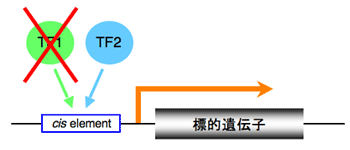
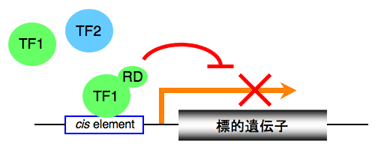
 図3
図3 図4
図4 図5
図5